
Abstract
Background
Chronic therapy with nitroglycerin (GTN) results in a rapid development of nitrate tolerance which is associated with an increased production of reactive oxygen species (ROS). According to recent studies, mitochondrial ROS formation and oxidative inactivation of the organic nitrate bioactivating enzyme mitochondrial aldehyde dehydrogenase (ALDH-2) play an important role for the development of nitrate and cross-tolerance.
Methods
Tolerance was induced by infusion of wild type (WT) and heterozygous manganese superoxide dismutase mice (Mn-SOD+/-) with ethanolic solution of GTN (12.5 μg/min/kg for 4 d). For comparison, the tolerance-free pentaerithrityl tetranitrate (PETN, 17.5 μg/min/kg for 4 d) was infused in DMSO. Vascular reactivity was measured by isometric tension studies of isolated aortic rings. ROS formation and aldehyde dehydrogenase (ALDH-2) activity was measured in isolated heart mitochondria.
Results
Chronic GTN infusion lead to impaired vascular responses to GTN and acetylcholine (ACh), increased the ROS formation in mitochondria and decreased ALDH-2 activity in Mn-SOD+/- mice. In contrast, PETN infusion did not increase mitochondrial ROS formation, did not decrease ALDH-2 activity and accordingly did not lead to tolerance and cross-tolerance in Mn-SOD+/- mice. PETN but not GTN increased heme oxygenase-1 mRNA in EA.hy 926 cells and bilirubin efficiently scavenged GTN-derived ROS.
Conclusion
Chronic GTN infusion stimulates mitochondrial ROS production which is an important mechanism leading to tolerance and cross-tolerance. The tetranitrate PETN is devoid of mitochondrial oxidative stress induction and according to the present animal study as well as numerous previous clinical studies can be used without limitations due to tolerance and cross-tolerance.
Similar content being viewed by others

Background
Although organic nitrates such as nitroglycerin (glyceryl trinitrate, GTN) have been used for over a century in the therapy of cardiovascular diseases such as stable and unstable angina [1] the underlying mechanisms of nitrate bioactivation and development of nitrate tolerance are not completely understood and are most likely multifactorial [2]. Previously, we found that 3 days of nitrate treatment doubled vascular superoxide (O2 -) production [3] which was also found in human bypass vessels from nitroglycerin (GTN) treated patients [4]. The crucial role of oxidative stress for the development of nitrate and cross-tolerance has been repeatedly demonstrated in numerous cell culture [5, 6], animal [7–10] and human studies [11–15]. Most beneficial effects of therapeutics on nitrate tolerance were based on antioxidative properties [16–18].
In 2002, the mitochondrial aldehyde dehydrogenase (ALDH-2), which is subject to an oxidative mechanism-based inactivation, has been identified as a GTN-metabolizing enzyme and a possible important component in the processes leading to tolerance [19]. This concept was supported by recent studies in ALDH-2 deficient mice (ALDH-2-/-) [20]. Our laboratory further substantiated this concept in an animal model of in vivo tolerance and extended previous observations by demonstrating that mitochondria are a major source of ROS formation in response to acute and chronic GTN challenges [21–23]. The importance of the ALDH-2 concept for clinical nitrate tolerance was proven by two independent clinical studies in Asian subjects with a point mutated, dysfunctional ALDH-2 [24, 25].
In the present study, we sought to characterize the role of mitochondria as a source of superoxide formation for the development of in vivo nitrate tolerance in response to GTN. We also used the alternative organic nitrate, PETN, which has been described to cause no tolerance, to see whether lack of tolerance induction is associated with a decrease in ROS formation. We also addressed the initial steps underlying PETN protective effects which have been described to be based on induction of heme oxygenase-1 and formation of the potent antioxidant bilirubin [26, 27].
Methods
Materials
PETN (with 80 % (w/w) lactose) was from Fluka (Buchs, Switzerland). For isometric tension studies, GTN was used from a Nitrolingual infusion solution (1 mg/ml) from G.Pohl-Boskamp (Hohenlockstedt, Germany). For induction of in vivo tolerance, GTN was used from a solution in ethanol (102 g/l) which was obtained from UNIKEM (Copenhagen, Denmark). L-012 (8-amino-5-chloro-7-phenylpyrido [3,4-d]pyridazine-1,4-(2H,3H)dione sodium salt) was purchased from Wako Pure Chemical Industries (Osaka, Japan). All other chemicals were of analytical grade and were obtained from Sigma-Aldrich, Fluka or Merck.
Animals and in vivo treatment
All animal treatment was in accordance with the Declaration of Helsinki and with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the U.S. National Institutes of Health and was granted by the Ethics Committee of the University Hospital Eppendorf and the University Hospital Mainz. We used male mice aged 7–10 months on a mixed genetic background (C57Bl/6 × 129/Ola). Experiments were performed with 14 wt and 14 Mn-SOD+/- mice. Mn-SOD+/- mice were generated according to a published procedure [28] in the laboratory of K. Scharffetter-Kochanek. The deficiency of the Mn-SOD activity was determined using a specific activity assay as described recently [28]. In vivo tolerance was induced by chronic infusion of mice with GTN in ethanol (25 μg/h, 60 nmol/min/kg for 4 d) by implanted micro-osmotic pumps (alzet, model 1007D, 0.5 μl/h for 7 d) from Durect Corp. (Cupertino, CA). Infusion of the solvent ethanol served as a control. We also infused mice with PETN in DMSO (35 μg/h, 60 nmol/min/kg) or the solvent alone. After this period, the animals were sacrificed and aorta as well as hearts were subjected to further analysis. The detailed protocol was recently published [23]. Wistar rats were either infused with ethanol (1 μl/h for 4 d) as the solvent control or GTN in ethanol (6.6 μg/kg/min for 4 d) to induce tolerance as previously published [16, 22]. In These Wistar rats we used micro-osmotic pumps (alzet, model 2001, 1 μl/h for 7 d) from Durect Corp. (Cupertino, CA).
Isometric tension studies
Vasodilator responses to GTN, PETN, ISDN and ACh were assessed with endothelium-intact isolated murine aortic rings mounted for isometric tension recordings in organ chambers upon preconstriction with prostaglandin F2α, as described previously [29]. It is important to note that the order of subsequent concentration-relaxation-curves was ACh, ISDN, GTN (in GTN in vivo treatment group) and ACh, PETN, GTN (in PETN in vivo treatment group). This order is important to explain the phenomenon of tachyphylaxis induced during tension studies in the organ baths.
Detection of oxidative stress in isolated heart mitochondria
Isolated mitochondria were prepared from mouse hearts according to a previously published protocol and ROS formation was detected by L-012 (100 μM) ECL as recently described [22, 23, 30]. Briefly, hearts from mice were homogenized and the 20,000 g pellet was resuspended in Tris buffer. Mitochondrial suspensions were diluted to a final protein concentration of 0.1 mg/ml in 0.5 ml of PBS buffer containing L-012 (100 μM). ROS production was detected after stimulation with succinate (4 mM final concentration). Mitochondria from rat heart were prepared by a similar procedure and ROS formation was detected by L-012 ECL as published [31]. Effect of bilirubin on ROS formation was tested at concentrations of 0.25–25 μM from a 1 mM stock in DMSO.
ALDH-2 dehydrogenase activity in isolated mouse heart mitochondria
The activity of ALDH-2 in isolated mitochondria was determined by measuring the conversion of 2-hydroxy-3-nitro-benzaldehyde to 2-hydroxy-3-nitro-benzoic acid using a modified published protocol [22]. The mitochondrial suspension was diluted to approximately 1 mg/ml protein in PBS and 2-hydroxy-3-nitro-benzaldehyde (100 μM) was added and the samples were incubated for 30 min at 37°C. 100 μl of each sample were subjected to HPLC analysis.
Effects of GTN and PETN on HO-1 mRNA expression in cultured EA.hy 926 cells
Human endothelial cells EA.hy 926 cells [32] were grown in Dulbecco's modified Eagle's medium (Sigma) with 10% fetal calf serum, 2 mM L-glutamine, 1 mM sodium pyruvate, 100 IU/ml penicillin, 100 μg/ml streptomycin, and 1× HAT (hypoxanthine, amethopterin/methotrexate, thymine) [33]. Confluent cells (6-well plates) were incubated with either DMSO (0.1 %), 10 μM PETN, 50 μM PETN or ethanol (0.1 %), 10 μM GTN, 50 μM GTN for 12 h. mRNA expression of HO-1 was analyzed with quantitative real-time RT.
Briefly, total RNA from EA.hy926 cells was isolated according to the manufacturer's protocol (RNeasy Mini Kit; Qiagen, Hilden, Germany). 0.5 μg of total RNA was used for real-time RT-PCR analysis with the QuantiTect™ Probe RT-PCR kit (Qiagen) in 25 μl reactions in a 96-well spectrofluorometric thermal cycler (iCycler, Bio-Rad, München, Germany). Real-time qRT-PCR was performed according to the manufacturer's recommendations using the oligonucleotides listed below (all from MWG-Biotech, Ebersberg, Germany).
HO-1
sense AGGCCAAGACTGCGTTCCT
antisense GGCTCTGGTCCTTGGTGTCAT
probe CTCAACATCCAGCTCTTTGAGGAGTTGCAG
GAPDH
sense CCCATGTTCGTCATGGGTGT
antisense TGGTCATGAGTCCTTCCACGATA
probe CTGCACCACCAACTGCTTAGCACCC
Each experimental reaction was performed in triplicate. All primer/probe sets had efficiencies of 100% (± 10%). The comparative Ct method was used for relative mRNA quantification [34]. Gene expression was normalized to the endogenous control, GAPDH mRNA, and the amount of target gene mRNA expression in each sample was expressed relative to that of control.
Statistical Analysis
Results are expressed as mean ± SEM. One-way ANOVA (with Bonferroni's or Dunn's correction for comparison of multiple means) was used for comparisons of vasodilator potency and efficacy, L-012-derived chemiluminescence, ALDH-2 activity as well as HO-1 mRNA expression. The EC50 value for each experiment was obtained by log-transformation. P values < 0.05 were considered significant. * indicates significance vs. solvent control.
Results
Effect of heterozygous deficiency in mitochondrial superoxide dismutase (Mn-SOD+/-) on vasodilator responses in response to organic nitrate treatment
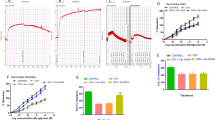
We here show for the first time data from isometric tension studies indicating that a partial (50 %) deficiency in Mn-SOD and accordingly increased basal mitochondrial ROS levels result in a more pronounced clinical tolerance. This phenomenon was characterized by impaired vasodilator potency of GTN and a significant right-shift of the concentration-relaxation curve (Figure 1A). Similar effects were observed for the concentration-relaxation curves of the endothelium-dependent vasodilator ACh as well as the nitrovasodilator isosorbide dinitrate (ISDN), indicating mild but not significant cross-tolerance to these compounds in GTN-treated Mn-SOD vessels (see Table 1). It is important to note that the GTN in vivo dose was adjusted as compared to previous studies to cause no severe tolerance in wild type control mice. Accordingly, GTN in vivo treatment of wild type mice with this low nitrate dose resulted in mild tolerance to GTN but obviously no cross-tolerance to ACh and ISDN (Table 1). In contrast, PETN in vivo treatment resulted in no clinical tolerance and we observed normal PETN and ACh potencies in vessels from wild type and Mn-SOD+/- mice (Figure 1B, Table 1). Nevertheless, the PETN concentration-response-curve which was performed prior to the GTN concentration-response-curve resulted in decreased potency of GTN (Table 1). This indicates that in vitro incubation with the concentration of PETN that caused 100 % relaxation (ED100 = 31.6 μM) induced in vitro tolerance (tachyphylaxis) to the finally applied nitrate GTN. The GTN potency in this subsequent concentration-relaxation-curve was shifted by more than one log as compared to normal GTN potency in mouse aorta (see wild type in Table 1).

Vasodilator responses of isolated aortic vessel segments upon chronic treatment of wild type and Mn-SOD +/- mice with GTN or PETN. (A) Concentration-relaxation curves for GTN (10-9 to 10-4.5 M) in vessels from wild type and Mn-SOD+/- mice upon treatment with ethanol or GTN in ethanol. The symbols are closed circles (ethanol infused wild type), closed triangles (GTN infused wild type), open circles (ethanol infused Mn-SOD+/-) and open triangles (GTN infused Mn-SOD+/-). Data are mean ± SEM of 7–10 independent experiments. (B) Concentration-relaxation curves for PETN (10-10 to 10-4.5 M) in vessels from wild type and Mn-SOD+/- mice upon treatment with DMSO or PETN in DMSO. The symbols are closed circles (DMSO infused wild type), closed triangles (PETN infused wild type), open circles (DMSO infused Mn-SOD+/-) and open triangles (PETN infused Mn-SOD+/-). Data are mean ± SEM of 6–8 independent experiments.
Effect of heterozygous deficiency in mitochondrial superoxide dismutase (Mn-SOD+/-) and bilirubin levels on mitochondrial oxidative stress in response to organic nitrate treatment
In accordance to the vascular responses, mitochondrial ROS was not increased but rather decreased in PETN-treated Mn-SOD+/- mice (Figure 2B). This was an important finding with respect to our recent observation that mitochondrial ROS levels were significantly increased upon GTN in vivo treatment of Mn-SOD+/- mice [23] which was reproduced with the present findings (Figure 2A). Since the applied GTN dose was low, wild type mitochondria showed no increased ROS formation in response to GTN in vivo treatment which could be best explained by antioxidant properties of the small amounts of the solvent ethanol. Ethanol in turn could interfere with GTN bioactivation by the formation of the natural substrate and competitive inhibitor of bioactivation acetaldehyde.

Mitochondrial ROS formation upon chronic treatment of wild type and Mn-SOD +/- mice with GTN or PETN and effects of bilirubin. (A) Mitochondrial ROS formation was stimulated with succinate (2 mM) and measured by L-012 (100 μM) ECL in hearts from wild type and Mn-SOD+/- mice upon treatment with ethanol or GTN in ethanol. Data are mean ± SEM of 4 independent experiments. * P < 0.05 vs. WT/EtOH and # P < 0.05 vs. MnSOD/EtOH. (B) Mitochondrial ROS formation was stimulated with succinate (2 mM) and measured by L-012 (100 μM) ECL in hearts from wild type and Mn-SOD+/- mice upon treatment with DMSO or PETN in DMSO. Data are mean ± SEM of 16 independent experiments. * P < 0.05 vs. WT/DMSO. (C) Bilirubin efficiently decreased mitochondrial ROS (L-012 ECL) in response to GTN in vivo treatment of Wistar rats. Data are mean ± SEM of 6–11 independent experiments. # P < 0.05 vs. control, * P < 0.05 vs. GTN in vivo group.
Mitochondrial ROS were increased by almost 50 % in response to GTN in vivo treatment of Wistar rats (Figure 2C). Bilirubin concentration-dependently decreased the chemiluminescence signal in mitochondria from GTN-treated Wistar rats indicating its potent scavenging properties for GTN-derived ROS (Figure 2C).
Effect of heterozygous deficiency in mitochondrial superoxide dismutase (Mn-SOD+/-) on mitochondrial ALDH-2 activity in response to organic nitrate treatment
GTN in vivo treatment decreased the redox-sensitive ALDH-2 activity in isolated heart mitochondria from Mn-SOD+/- mice (30.4 ± 1.5 in ethanol group vs. 21.4 ± 1.5 in GTN group, n = 25–27, p < 0.001) whereas PETN slightly (although not significantly) increased the ALDH-2 activity (39.8 ± 3.3 in DMSO group vs. 42.3 ± 4.4 in PETN group, n = 9, p = 0.647).
Effects of GTN and PETN on HO-1 mRNA expression in cultured EA.hy 926 cells
PETN concentration-dependently increased the HO-1 mRNA levels in EA.hy 926 cells resulting in a significant increase at 50 μM (Figure 3). In contrast, GTN only induced a minor increase in HO-1 mRNA which did not reach significance (Figure 3). Similar differences have previously been demonstrated between the PETN trinitrate metabolite PETriN and ISDN [26, 27].

HO-1 mRNA expression in EA.hy 926 cells in response to PETN or GTN treatment. Confluent cells were incubated with the solvent (DMSO or ethanol) or PETN as well as GTN for 12 h. PETN but not GTN resulted in a significant increase in HO-1 mRNA. Data are mean ± SEM of at least 3 independent experiments. * P < 0.05 vs. control.
Discussion
Nitrate tolerance and the "oxidative stress concept"
Previous studies have demonstrated that nitrate tolerance in response to GTN in vivo treatment is a multifactorial phenomenon [2]. The "oxidative stress concept" in the setting of nitrate tolerance was established by our group [3] and refined during the last couple of years [4, 23]. In essence, the concept consists of increased superoxide formation in response to nitrate treatment which decreases NO bioavailability, leads to peroxynitrite formation [8], NOS uncoupling [35] and impairs NO/cGMP signaling [4]. Moreover, oxidative inhibition of prostacyclin synthase [18] as well as mitochondrial ALDH activity [22] may present other key events in the development of nitrate tolerance. In the present study we demonstrate that low dose GTN for 4 d causes tolerance (indicated by a significant right-shift in GTN concentration-relaxation-curve) in Mn-SOD+/- mice but has only minor effects on GTN potency and efficacy in wild type mice (Figure 1A). Also mild cross-tolerance to ACh and ISDN was observed (Table 1). In contrast, PETN which has been reported to possess protective effects, caused no tolerance (Figure 1B and Table 1).
Inhibition of ALDH-2 and mitochondrial ROS formation
Recently, we have identified the mitochondria as a major source of oxidative stress in tolerant animals [16, 21, 22]. These results are in accordance with the observation that the mitochondrial ALDH provides an important bioactivation pathway for GTN [19, 20, 24, 25] and is redox-sensitive [36, 37] (see scheme in Figure 4). These facts provide a new link between oxidative stress and impaired GTN bioactivation and accordingly development of tolerance: Mitochondrial ROS oxidatively inactivate the ALDH-2. The involvement of mitochondria in the development of nitrate tolerance was previously postulated by the "thiol concept" of Needleman and coworkers [38, 39]. An interesting link between mitochondrial oxidative stress triggered ALDH-2 inhibition and activation of vascular NADPH oxidases could be the accumulation of toxic aldehydes (as observed in ALDH-2-/- mice). It was reported that low concentrations of 9-hydroxynonenal may stimulate whereas high concentrations may inhibit NADPH oxidase activity and/or expression [40]. In the present study we could demonstrate that GTN treatment resulted in an increase in mitochondrial ROS and a decrease in ALDH-2 activity in the Mn-SOD+/- mice whereas PETN did not significantly alter these parameters (Figure 2 and Results).

Proposed mechanism for organic nitrate bioactivation, induction of oxidative stress and protective effects. Highly potent organic nitrate (tri- and tetranitrates) are bioactivated by mitochondrial ALDH. The bioactivation (reduction cycle) requires two thiol groups at the active site of the enzyme which are oxidized to the disulfide during the conversion which yields the denitrated metabolite and an NOx species that is similar to NO. Enzymatic activity is restored by reduced lipoic acid/lipoamide which is recycled by lipoamide reductase (LAR), thioredoxin/thioredoxin reductase system (Trx/TrxR) or glutathione/glutathionereductase system (GSH/GR). GTN leads to mechanism-based inactivation of the enzyme but also triggers mitochondrial oxidative stress which may directly inactivate mtALDH, deplete dihydrolipoic acid or interfere with its reductases. In contrast, PETN induces HO-1 which by breakdown of porphyrins yields the potent antioxidant bilirubin, the anti-inflammatory compound CO and induces ferritin, another protective enzyme which decreases free iron and prevents Fenton-type reactions.
Diversity of organic nitrates with respect to induction of oxidative stress and clinical tolerance
The tetranitrate PETN was previously shown to cause no induction of clinical tolerance and vascular oxidative stress [12, 14, 17]. The beneficial properties of PETN were explained by induction of the antioxidative proteins ferritin and HO-1 [26, 27] which may prevent oxidative stress and protect the vasculature from oxidative damage [2] and thereby mimic the antioxidant principle of compounds such as hydralazine [16]. HO-1 has been demonstrated to be a major protective and antioxidative principle in numerous therapeutic interventions [41, 42]. The underlying mechanism of this protection is thought to be based on the breakdown of porphyrins to yield the potent antioxidant bilirubin [43–45] and the anti-inflammatory compound carbon monoxide [46, 47] which is a weak activator of soluble guanylyl cyclase [48]. This is of special interest since HO-1 was reported to be localized within mitochondria [49] and thereby could directly affect nitrate-induced mitochondrial ROS formation and protect ALDH-2 from oxidative inactivation (see scheme in Figure 4).
There is growing body of evidence that ALDH-2 only bioactivates tri- and tetranitrates which show high potency in tension studies. During reduction of nitrates the dithiol groups at the active site of the enzyme form a disulfide bridge causing its inactivation. Dihydrolipoic acid is able to restore enzymatic activity (Daiber et al., unpublished observations) but needs reduction by appropriate enzymes. It is thought that GTN induces mitochondrial ROS formation which may contribute to oxidative inhibition of ALDH-2 activity and depletion of reduced thiols (such as dihydrolipoic acid) [38] thereby disrupting the physiological catalytic cycle (Figure 4). According to numerous reports in the literature PETN displays potent antioxidative properties which are probably based on the afore mentioned formation of bilirubin, CO and ferritin (Figure 4). In the present study we could demonstrate that bilirubin efficiently decreases mitochondrial oxidative stress in response to GTN treatment (Figure 2C). Moreover, we could reproduce previous results of Oberle et al. on PETN-triggered HO-1 induction and more importantly demonstrate that this inducing effect is not shared by GTN (Figure 3). This, however, could provide an explanation for the differences in tolerance induction by both organic nitrates.
One would expect that all NO donors induce HO-1 since this gene is heavily regulated by NO. However, previous publications have demonstrated that HO-1 inducing capacity is not shared by all NO donors [50]. It is unclear why ISDN and GTN do not induce HO-1. Especially, since HO-1 is regulated by at least 3 important transcription factors (NFκB, AP-1, Nrf2/Keap1) and induced by a huge number of different stimuli [51] providing many regulatory sites, which could explain the different properties of ISDN and GTN versus PETN. At this stage we have no hypothesis to explain these differences in vitro.
Conclusion
The results obtained in Mn-SOD+/- mice clearly indicate that mitochondrial ROS play an important role for the development/maintenance of nitrate tolerance. The results obtained with PETN indicate that organic nitrates differ markedly and that protective antioxidative properties (mediated by HO-1 and derived bilirubin) may compensate for harmful induction of oxidative stress sources.
Abbreviations
- ACh :
-
acetylcholine
- ALDH-2 :
-
mitochondrial aldehyde dehydrogenase
- ECL :
-
enhanced chemiluminescence
- GTN :
-
glyceryl trinitrate (nitroglycerin)
- HO-1 :
-
heme oxygenase-1
- ISDN :
-
isosorbide dinitrate
- L-012 :
-
8-amino-5-chloro-7-phenylpyrido [3,4-d]pyridazine-1,4-(2H,3H)dione sodium salt
- Mn-SOD :
-
manganese superoxide dismutase (mitochondrial isoform)
- Mn-SOD :
-
+/-:heterozygous Mn-SOD deficiency
- PETN :
-
pentaerithrityl tetranitrate
- ROS :
-
reactive oxygen species.
References
Abrams J: The role of nitrates in coronary heart disease. Arch Intern Med. 1995, 155 (4): 357-364. 10.1001/archinte.155.4.357.
Munzel T, Daiber A, Mulsch A: Explaining the phenomenon of nitrate tolerance. Circ Res. 2005, 97 (7): 618-628. 10.1161/01.RES.0000184694.03262.6d.
Munzel T, Sayegh H, Freeman BA, Tarpey MM, Harrison DG: Evidence for enhanced vascular superoxide anion production in nitrate tolerance. A novel mechanism underlying tolerance and cross-tolerance. J Clin Invest. 1995, 95 (1): 187-194.
Schulz E, Tsilimingas N, Rinze R, Reiter B, Wendt M, Oelze M, Woelken-Weckmuller S, Walter U, Reichenspurner H, Meinertz T, Munzel T: Functional and biochemical analysis of endothelial (dys)function and NO/cGMP signaling in human blood vessels with and without nitroglycerin pretreatment. Circulation. 2002, 105 (10): 1170-1175. 10.1161/hc1002.105186.
Abou-Mohamed G, Johnson JA, Jin L, El-Remessy AB, Do K, Kaesemeyer WH, Caldwell RB, Caldwell RW: Roles of superoxide, peroxynitrite, and protein kinase C in the development of tolerance to nitroglycerin. J Pharmacol Exp Ther. 2004, 308 (1): 289-299. 10.1124/jpet.103.056119.
Abou-Mohamed G, Kaesemeyer WH, Caldwell RB, Caldwell RW: Role of L-arginine in the vascular actions and development of tolerance to nitroglycerin. Br J Pharmacol. 2000, 130 (2): 211-218. 10.1038/sj.bjp.0703293.
Munzel T, Kurz S, Rajagopalan S, Thoenes M, Berrington WR, Thompson JA, Freeman BA, Harrison DG: Hydralazine prevents nitroglycerin tolerance by inhibiting activation of a membrane-bound NADH oxidase. A new action for an old drug. J Clin Invest. 1996, 98 (6): 1465-1470.
Warnholtz A, Mollnau H, Heitzer T, Kontush A, Moller-Bertram T, Lavall D, Giaid A, Beisiegel U, Marklund SL, Walter U, Meinertz T, Munzel T: Adverse effects of nitroglycerin treatment on endothelial function, vascular nitrotyrosine levels and cGMP-dependent protein kinase activity in hyperlipidemic Watanabe rabbits. J Am Coll Cardiol. 2002, 40 (7): 1356-1363. 10.1016/S0735-1097(02)02133-2.
Bassenge E, Fink N, Skatchkov M, Fink B: Dietary supplement with vitamin C prevents nitrate tolerance. J Clin Invest. 1998, 102 (1): 67-71.
Fink B, Schwemmer M, Fink N, Bassenge E: Tolerance to nitrates with enhanced radical formation suppressed by carvedilol. J Cardiovasc Pharmacol. 1999, 34 (6): 800-805. 10.1097/00005344-199912000-00005.
Munzel T, Hayoz D, Hornig B, Kurz S, Just H, Zelis B, Brunner HR, Drexler H: [Increased vascular sensitivity to nitroglycerin in patients with hypercholesterolemia and peripheral endothelial dysfunction]. Deutsche Medizinische Wochenschrift. 1994, 119 (31-32): 1065-1070.
Jurt U, Gori T, Ravandi A, Babaei S, Zeman P, Parker JD: Differential effects of pentaerythritol tetranitrate and nitroglycerin on the development of tolerance and evidence of lipid peroxidation: a human in vivo study. J Am Coll Cardiol. 2001, 38 (3): 854-859. 10.1016/S0735-1097(01)01414-0.
Gori T, Burstein JM, Ahmed S, Miner SE, Al-Hesayen A, Kelly S, Parker JD: Folic acid prevents nitroglycerin-induced nitric oxide synthase dysfunction and nitrate tolerance: a human in vivo study. Circulation. 2001, 104 (10): 1119-1123.
Keimer R, Stutzer FK, Tsikas D, Troost R, Gutzki FM, Frolich JC: Lack of oxidative stress during sustained therapy with isosorbide dinitrate and pentaerythrityl tetranitrate in healthy humans: a randomized, double-blind crossover study. J Cardiovasc Pharmacol. 2003, 41 (2): 284-292. 10.1097/00005344-200302000-00018.
Skatchkov M, Larina LL, Larin AA, Fink N, Bassenge E: Urinary NItrotyrosine Content as a Marker of Peroxynitrite-induced Tolerance to Organic NItrates. J Cardiovasc Pharmacol Ther. 1997, 2 (2): 85-96.
Daiber A, Oelze M, Coldewey M, Kaiser K, Huth C, Schildknecht S, Bachschmid M, Nazirisadeh Y, Ullrich V, Mulsch A, Munzel T, Tsilimingas N: Hydralazine is a powerful inhibitor of peroxynitrite formation as a possible explanation for its beneficial effects on prognosis in patients with congestive heart failure. Biochem Biophys Res Commun. 2005, 338 (4): 1865-1874. 10.1016/j.bbrc.2005.10.106.
Schwemmer M, Bassenge E: New approaches to overcome tolerance to nitrates. Cardiovasc Drugs Ther. 2003, 17 (2): 159-173. 10.1023/A:1025343919959.
Hink U, Oelze M, Kolb P, Bachschmid M, Zou MH, Daiber A, Mollnau H, August M, Baldus S, Tsilimingas N, Walter U, Ullrich V, Munzel T: Role for peroxynitrite in the inhibition of prostacyclin synthase in nitrate tolerance. J Am Coll Cardiol. 2003, 42 (10): 1826-1834. 10.1016/j.jacc.2003.07.009.
Chen Z, Zhang J, Stamler JS: Identification of the enzymatic mechanism of nitroglycerin bioactivation. Proc Natl Acad Sci U S A. 2002, 99 (12): 8306-8311. 10.1073/pnas.122225199.
Chen Z, Foster MW, Zhang J, Mao L, Rockman HA, Kawamoto T, Kitagawa K, Nakayama KI, Hess DT, Stamler JS: An essential role for mitochondrial aldehyde dehydrogenase in nitroglycerin bioactivation. Proc Natl Acad Sci U S A. 2005, 102 (34): 12159-12164. 10.1073/pnas.0503723102.
Sydow K, Daiber A, Oelze M, Chen Z, August M, Wendt M, Ullrich V, Mulsch A, Schulz E, Keaney JF, Stamler JS, Munzel T: Central role of mitochondrial aldehyde dehydrogenase and reactive oxygen species in nitroglycerin tolerance and cross-tolerance. J Clin Invest. 2004, 113 (3): 482-489. 10.1172/JCI200419267.
Daiber A, Oelze M, Coldewey M, Bachschmid M, Wenzel P, Sydow K, Wendt M, Kleschyov AL, Stalleicken D, Ullrich V, Mulsch A, Munzel T: Oxidative stress and mitochondrial aldehyde dehydrogenase activity: a comparison of pentaerythritol tetranitrate with other organic nitrates. Mol Pharmacol. 2004, 66 (6): 1372-1382. 10.1124/mol.104.002600.
Daiber A, Oelze M, Sulyok S, Coldewey M, Schulz E, Treiber N, Hink U, Mulsch A, Scharffetter-Kochanek K, Munzel T: Heterozygous Deficiency of Manganese Superoxide Dismutase in Mice (Mn-SOD+/-): A Novel Approach to Assess the Role of Oxidative Stress for the Development of Nitrate Tolerance. Mol Pharmacol. 2005, 68 (3): 579-588.
Mackenzie IS, Maki-Petaja KM, McEniery CM, Bao YP, Wallace SM, Cheriyan J, Monteith S, Brown MJ, Wilkinson IB: Aldehyde dehydrogenase 2 plays a role in the bioactivation of nitroglycerin in humans. Arterioscler Thromb Vasc Biol. 2005, 25 (9): 1891-1895. 10.1161/01.ATV.0000179599.71086.89.
Li Y, Zhang D, Jin W, Shao C, Yan P, Xu C, Sheng H, Liu Y, Yu J, Xie Y, Zhao Y, Lu D, Nebert DW, Harrison DC, Huang W, Jin L: Mitochondrial aldehyde dehydrogenase-2 (ALDH2) Glu504Lys polymorphism contributes to the variation in efficacy of sublingual nitroglycerin. J Clin Invest. 2006, 116 (2): 506-511. 10.1172/JCI26564.
Oberle S, Abate A, Grosser N, Hemmerle A, Vreman HJ, Dennery PA, Schneider HT, Stalleicken D, Schroder H: Endothelial protection by pentaerithrityl trinitrate: bilirubin and carbon monoxide as possible mediators. Exp Biol Med (Maywood). 2003, 228 (5): 529-534.
Oberle S, Schwartz P, Abate A, Schroder H: The antioxidant defense protein ferritin is a novel and specific target for pentaerithrityl tetranitrate in endothelial cells. Biochem Biophys Res Commun. 1999, 261 (1): 28-34. 10.1006/bbrc.1999.0941.
Strassburger M, Bloch W, Sulyok S, Schuller J, Keist AF, Schmidt A, Wenk J, Peters T, Wlaschek M, Krieg T, Hafner M, Kumin A, Werner S, Muller W, Scharffetter-Kochanek K: Heterozygous deficiency of manganese superoxide dismutase results in severe lipid peroxidation and spontaneous apoptosis in murine myocardium in vivo. Free Radic Biol Med. 2005, 38 (11): 1458-1470. 10.1016/j.freeradbiomed.2005.02.009.
Daiber A, Oelze M, Sulyok S, Coldewey M, Schulz E, Treiber N, Hink U, Mulsch A, Scharffetter-Kochanek K, Munzel T: Heterozygous deficiency of manganese superoxide dismutase in mice (Mn-SOD+/-): a novel approach to assess the role of oxidative stress for the development of nitrate tolerance. Mol Pharmacol. 2005, 68 (3): 579-588.
Munzel T, Giaid A, Kurz S, Stewart DJ, Harrison DG: Evidence for a role of endothelin 1 and protein kinase C in nitroglycerin tolerance. Proc Natl Acad Sci U S A. 1995, 92 (11): 5244-5248. 10.1073/pnas.92.11.5244.
Raha S, McEachern GE, Myint AT, Robinson BH: Superoxides from mitochondrial complex III: the role of manganese superoxide dismutase. Free Radic Biol Med. 2000, 29 (2): 170-180. 10.1016/S0891-5849(00)00338-5.
Daiber A, Oelze M, August M, Wendt M, Sydow K, Wieboldt H, Kleschyov AL, Munzel T: Detection of superoxide and peroxynitrite in model systems and mitochondria by the luminol analogue L-012. Free Radic Res. 2004, 38 (3): 259-269. 10.1080/10715760410001659773.
Edgell CJ, McDonald CC, Graham JB: Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc Natl Acad Sci U S A. 1983, 80 (12): 3734-3737. 10.1073/pnas.80.12.3734.
Li H, Oehrlein SA, Wallerath T, Ihrig-Biedert I, Wohlfart P, Ulshofer T, Jessen T, Herget T, Forstermann U, Kleinert H: Activation of protein kinase C alpha and/or epsilon enhances transcription of the human endothelial nitric oxide synthase gene. Mol Pharmacol. 1998, 53 (4): 630-637.
Livak KJ, Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001, 25 (4): 402-408. 10.1006/meth.2001.1262.
Munzel T, Li H, Mollnau H, Hink U, Matheis E, Hartmann M, Oelze M, Skatchkov M, Warnholtz A, Duncker L, Meinertz T, Forstermann U: Effects of long-term nitroglycerin treatment on endothelial nitric oxide synthase (NOS III) gene expression, NOS III-mediated superoxide production, and vascular NO bioavailability. Circ Res. 2000, 86 (1): E7-E12.
Senior DJ, Tsai CS: Esterase activity of high-Km aldehyde dehydrogenase from rat liver mitochondria. Biochem Cell Biol. 1990, 68 (4): 758-763.
Tsai CS, Senior DJ: Chemical studies of high-Km aldehyde dehydrogenase from rat liver mitochondria. Biochem Cell Biol. 1991, 69 (2-3): 193-197.
Needleman P, Hunter FE: Effects of organic nitrates on mitochondrial respiration and swelling: possible correlations with the mechanism of pharmacologic action. Mol Pharmacol. 1966, 2 (2): 134-143.
Needleman P, Blehm DJ, Rotskoff KS: Relationship between glutathione-dependent dentration and the vasodilator effectiveness of organic nitrates. J Pharmacol Exp Ther. 1969, 165 (2): 286-288.
Di Mauro C, Cavalli G, Amprimo MC, Paradisi L, Scano G, Curzio M, Dianzani MU: Influence of 4-hydroxynonenal on chemiluminescence production by unstimulated and opsonized zymosan-stimulated human neutrophils. Cell Biochem Funct. 1990, 8 (3): 147-155. 10.1002/cbf.290080304.
Grosser N, Hemmerle A, Berndt G, Erdmann K, Hinkelmann U, Schurger S, Wijayanti N, Immenschuh S, Schroder H: The antioxidant defense protein heme oxygenase 1 is a novel target for statins in endothelial cells. Free Radic Biol Med. 2004, 37 (12): 2064-2071. 10.1016/j.freeradbiomed.2004.09.009.
Immenschuh S, Schroder H: Heme oxygenase-1 and cardiovascular disease. Histol Histopathol. 2006, 21 (6): 679-685.
Yamaguchi T, Terakado M, Horio F, Aoki K, Tanaka M, Nakajima H: Role of bilirubin as an antioxidant in an ischemia-reperfusion of rat liver and induction of heme oxygenase. Biochem Biophys Res Commun. 1996, 223 (1): 129-135. 10.1006/bbrc.1996.0857.
Minetti M, Mallozzi C, Di Stasi AM, Pietraforte D: Bilirubin is an effective antioxidant of peroxynitrite-mediated protein oxidation in human blood plasma. Arch Biochem Biophys. 1998, 352 (2): 165-174. 10.1006/abbi.1998.0584.
Stocker R, Yamamoto Y, McDonagh AF, Glazer AN, Ames BN: Bilirubin is an antioxidant of possible physiological importance. Science. 1987, 235 (4792): 1043-1046. 10.1126/science.3029864.
Otterbein LE, Bach FH, Alam J, Soares M, Tao Lu H, Wysk M, Davis RJ, Flavell RA, Choi AM: Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat Med. 2000, 6 (4): 422-428. 10.1038/74680.
Morita T: Heme oxygenase and atherosclerosis. Arterioscler Thromb Vasc Biol. 2005, 25 (9): 1786-1795. 10.1161/01.ATV.0000178169.95781.49.
Brune B, Ullrich V: Inhibition of platelet aggregation by carbon monoxide is mediated by activation of guanylate cyclase. Mol Pharmacol. 1987, 32 (4): 497-504.
Converso DP, Taille C, Carreras MC, Jaitovich A, Poderoso JJ, Boczkowski J: HO-1 is located in liver mitochondria and modulates mitochondrial heme content and metabolism. Faseb J. 2006, 20 (8): 1236-1238. 10.1096/fj.05-4204fje.
Grosser N, Schroder H: [Therapy with NO donors-antiatherogenic and antioxidant actions]. Herz. 2004, 29 (1): 116-122. 10.1007/s00059-004-2530-3.
Prawan A, Kundu JK, Surh YJ: Molecular basis of heme oxygenase-1 induction: implications for chemoprevention and chemoprotection. Antioxid Redox Signal. 2005, 7 (11-12): 1688-1703. 10.1089/ars.2005.7.1688.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2261/6/44/prepub
Acknowledgements
We thank Jörg Schreiner for expert technical assistance. The present work was supported by a vascular biology grant from Actavis Deutschland GmbH (T.M.) as well as continuous funding by the German Research Foundation (DFG) (SFB 553 – C17 to T.M. and A.D.) and by the European Vascular Genomic Network, a Network of Excellence supported by the European Community's sixth Framework Program (Workpackages 1–3).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
DS is an employee of Actavis Deutschland GmbH and TM received vascular research grants from Actavis Deutschland GmbH.
Authors' contributions
1) HM: principal investigator, has made substantial contributions to conception and design of the study
2) PW: second principal investigator, has made substantial contributions to conception and design of the study
3) MO: performed animal surgery
4) NT: performed breeding of MnSOD+/- mice
5) AP: performed mRNA measurements
6) ES: cultured EA.hy 926 cells
7) SS: performed incubations of EA.hy 926 cells
8) KR: was involved in animal care and critically revised the manuscript
9) DS: was involved in drafting and interpretation
10) KSK: was involved in drafting and interpretation
11) HK: was involved in drafting, interpretation and mRNA measurement
12) TM: was involved in drafting and interpretation and has made substantial contributions to conception and design of the study of data
13) AD: performed ROS and ALDH-2 measurements, was involved in drafting and interpretation and has made substantial contributions to conception and design of the study of data
Declaration: All authors read and approved the final manuscript
Hanke Mollnau, Philip Wenzel contributed equally to this work.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Mollnau, H., Wenzel, P., Oelze, M. et al. Mitochondrial oxidative stress and nitrate tolerance – comparison of nitroglycerin and pentaerithrityl tetranitrate in Mn-SOD+/-mice. BMC Cardiovasc Disord 6, 44 (2006). https://doi.org/10.1186/1471-2261-6-44
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2261-6-44